Hey everyone! Just wanted to pop in and introduce myself as a new contributor to this blog. Currently, I’m in my second year of residency and will therefore be subjecting you all to a whole lot of gen path! I’ll do my best to add interesting papers and odd tidbits that aren’t related to phase I, too.
Hi All, Thank you all for visiting. I am going to expire the site as of Aug 26 this year. I have not been able to contribute much, and it is time to move on.
This is ambitious, and I’m already off track, but as someone with a love for lists I figured I would at least attempt to plan out how I’m going to prepare for phase II (which I take in August 2020).
As far as prepping for histo, I’m going to do about 5 years of Wednesday slide conference slides. This evens out to about 10-12 a week (about 10 months to complete). The last two months will be for reviewing slides and going through Tinker’s list.
I already have summaries of JKP and McGavin, so I’ll be reading those texts along with the summaries. Each volume of JKP will be finished in about 3 months. McGavin I will summarize weekly on the blog (fingers crossed!) and will finish that…eventually.
All Vet Path summaries will be done by January, then I’ll organize them by species and just try to cram as much as I can later on next year (April-July). The other journals I’m planning on just cherry picking relevant articles to read in my down time.
Each other book on the additional sources list I have given myself one month to summarize, a lot of them I have written summaries for already, so hopefully this wont be too painful.
If anyone wants any of the materials I have summarized, please comment below with your email address and I’ll send them your way. Comments, great study tips, and practice test ideas are welcome!
I’m going to summarize the finer points of intrinsic and extrinsic apoptosis. As always, if you have comments, concerns, or corrections, please comment below!
Intrinsic and extrinsic apoptosis differ in how they are initiated and how they go about activating executioner caspases (caspase 3 and 6 are the major players). Intrinsic is initiated by lots of things including ER stress, decreased growth factors, and genetic damage, among others. Meanwhile, extrinsic is initiated by binding of Fas-FasL forming a Fas associated death domain (FADD) on the cell undergoing apoptosis.
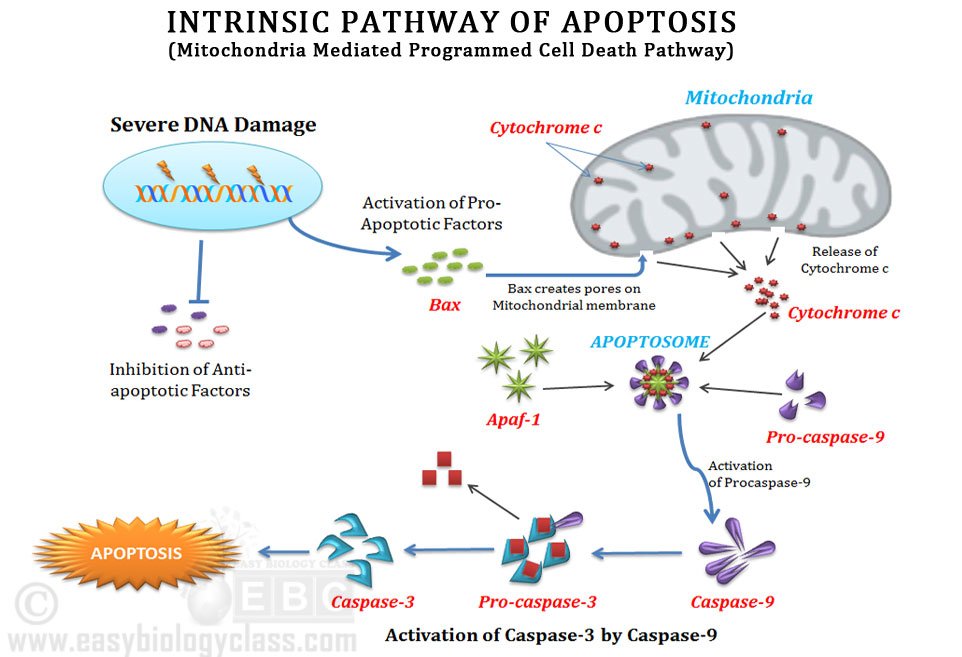
Intrinsic apoptosis starts with some mechanism OTHER than Fas-FasL binding, things like DNA damage, ER stress, decreased growth factors, etc. Sensors of apoptosis, the BH3 proteins (Bim, Bid, PUMA, NOXA, etc) are upregulated telling the cell to undergo apoptosis. If the cell has undergone DNA damage, the BH3 protein PUMA (p53 upregulated modulator of apoptosis) may have increased expression levels due to increased p53 levels.
Once the cell has committed (so to speak) to undergo intrinsic apoptosis, the mitochondrial membrane proteins that are anti-apoptotic are replaced with pro-apoptotic proteins (Bcl-2/Bcl-x/Mcl-1 –> Bax/Bak). The way I remember that is the proteins without an “a” are anti-apoptotic, the proteins with an “a” are pro-apoptotic. Bax and Bak allow the mitochondria to leak proteins, including cytochrome C. Cytochrome C binds to APAF-1 (apoptosis-activating factor-1) which also binds to caspase-9, forming the apoptosome. This activates caspase-9 allowing to activation of the executioner caspases and DNAse activity downstream.
There are inhibitors of apoptosis (IAPs- including IAP, IAP1, cIAP2, XIAP, I’m sure there’s more) that are active within the cytosol. When Bak/Bax create pores in the mitochrondrial membrane, Smac/Diablo is released along with cytochrome C. Smac/Diablo function to block IAPs (inhibiting the inhibitors = pro-apoptotic).
Extrinsic apoptosis is initiated by the binding of Fas (CD95) to FasL (but binding to TRAIL and the TNFα receptor TNFR1 can also induce the extrinsic pathway of apoptosis). When any of those receptors are activated they create a FADD (or TRADD)- Fas-associated death domain, where three receptors are brought close together. A FADD/TRADD will bring together pro-caspase 8 molecules close together, which is autocatalytic for cleavage into active caspase 8 (the way I remember that caspase 8 is with extrinsic apoptosis is that eight and extrinsic both start with “e”). Caspase 8 then activates executioner caspases 3 and 6 which leads to apoptosis.
The protein FLIP (aka cFLIP, CFLAR) binds to, but does not activate, pro-caspase 8, making it an anti-apoptotic protein.
There can be cross over between the two pathways. When Bid (BH3 protein) is activated, in Fas signaling induced apoptosis (extrinsic) it can activate the incorporation of Bax/Bak into the mitochondrial membrane and subsequent cytochrome c release (leading to intrinsic apoptosis).
I’m not great at cramming, so my study plan probably wont work for some of you. There are people that only study for about a month before the test and do fine, I (unfortunately for my time) am not one of those people. Here’s what I planned out for Phase I-
I know everyone’s institution does Gen Path prep differently. At ours, by the time you’re ready to prep for boards you have a summary of Robbins and Cotran and Zachary and McGavin. I was also gifted a really useful resource, that hopefully you’ve heard of- the Famke Aeffner and Cynthia de Vries 2013 compilation of R&C (8th ed) and McGavin (5th ed). Although it’s getting a little old, it’s largely still relevant (again, comment below if you’re interested in a copy).
So I reorganized my textbook summaries to follow the Aeffner and de Vries summary and planned to start studying six months before the test (Mid-Sept). There are 8 sections in those notes, so three weeks a section is 24 weeks (of the 26). Plus starting one month before the test going through each section again in three days. That way I have about a week before the test to go through the things that are not making sense and read review articles.
Here are some links to great practices tests to utilize:
Yikes. Phase 1 is upon us. Less than a month to go, how are you all feeling for it? Personally, I’m not feeling great…but I do have a pretty detailed study plan (see upcoming post for a boards prep plan!) that’s helping to ease my worries. I’m going to be going through some of the important pathways and try to break them down into small nuggets of (hopefully) less painful gen path content.
First up, Notch signaling.
(Taken from Zacharioudaki E and SJ Bray. 2014. Tools and methods for studying Notch signaling in Drosophila melanogaster. Methods 68(1):173-182)
So the Notch gene is a highly conserved, it’s important not only during embryogenesis (developmental decisions including differentiation of tissue types) but in life after-birth too (regulating homeostasis, cancer development, immunity). Essentially, the Notch pathway has three over-arching functions (1) lateral inhibition, (2) lateral induction, and (3) lineage decisions. Lateral inhibition is when a bunch of multipotent cells that are adjacent develop a hierarchy of function so they do not all become the same cell type. On the other hand, lateral induction is when cells of one population determines the outcome of another cell population. Lastly, lineage decisions occur during asymmetric division. This allows daughter cells to develop into different populations of cells depending on Notch pathway modulators and expression.
Although this pathway has many functions, there actually aren’t any second messengers and the mechanism is pretty straightforward (thank goodness). Essentially, a ligand binds to the Notch receptor (a heterodimeric, single-pass transmembrane protein) which releases cleaved Notch into the cytoplasm. The cleaved Notch is then translocated to the nucleus where it targets certain transcription genes (via interaction with CBF1/Suppressor of Hairless/Lag-1, depending on the species/scenario we’re talking about). This was a description of canonical Notch signaling, non-canonical exists too and has different ligands and end-points than canonical signaling.
The cool thing about not having second messengers is that there is an important relationship between the amount of ligand that binds Notch and the response the cells undergoes.
As far as test questions seem to go, most of the focus (at least on practice questions) seems to be on the role Notch plays in regulating angiogenesis. So we’ll dive into that.
Both Notch (binding to the Dll4 ligand) and VEGF-A (binding to VEGF2 Receptor, mostly) are critical for angiogenesis and have opposing effects. Notch/Dll4 has inhibitory effects, limiting sprouting of vessels (even to the point of down regulating VEGF2R expression). VEGF expression actually induces Dll4 expression (negative feedback!) which helps to ensure that supporting cells and endothelial cells don’t occlude the new vessels being created.
Typically, VEGF expression is higher in the leading cell than in the cells closer to the sprout (where it’s coming from the parent vessel). There are a lot more details in Zachary and McGavin on the nitty gritty of angiogenesis (EPCs, BMP, PDGF, oh my!), but the information above is the basics on Notch/Dll4 involvement in the process.
Questions, comments, and corrections are welcome in the comments!
References:
Andersson ER, R Sandberg, and U Lendahl. 2011. Notch signaling: simplicity in design, verstaliiy in function. Development. 138:3593-3612
Siekmann AF and ND Lawson. 2007. Notch signalling and the regulation of angiogenesis. Cell Adhesion and Migration. 1:2, 104-106








You must be logged in to post a comment.